|
|
Эволюция растений. Смена флор. Теория преадаптации. Верхнее-пермская и ранне-девонская флоры |
|
Любой современный объективно написанный курс по тому или иному теоретическому разделу биологии больше напоминает задачник, чем свод твердо установленных положений. Мы можем перечислить то, что нам дано, но когда надо ответить на то, что спрашивается, большей частью мы становимся в тупик. В этот удивительный задачник природа забыла вставить лист с ответами. И чем важнее вопрос, тем дальше мы находимся от его решения. Говоря словами П. Эр лиха и Р. Холма, "основной вопрос остается без ответа: почему в ходе эволюции ДНК создала для своего собственного воспроизведения трубкозубов и людей, тогда как бактерии и другие простые организмы, казалось бы, могут служить не хуже для этой цели?". Мы не будем рассуждать на эту вечную тему, в которой есть что‑то сродни вопросу о смысле жизни. Реми Шовсп справедливо пишет: "От того, что мы будем задавать себе подобные неразрешимые вопросы, наука, право же, ничего не приобретет". А поэтому постараемся меньше спрашивать "почему", а больше уделим внимания вопросу "как?".
Кому отвечать на вопросы
Как сменялись во времени группы растений? Какие события определили эти смены! Подобные вопросы исследователи задают себе уже давно. Казалось бы, кто, как не палеоботаники, должны на них ответить? Ведь именно в их распоряжении находятся соответствующие "документы". Действительно, имеющаяся у палеоботаников информация огромна. Следуя из глубин геологического прошлого к нашим дням, мы видим, что эпоха господства древних псилофитов, прапапоротников и других споровых растений сменилась эпохой голосеменных. На последнем этапе истории растений на передний план вышли покрытосеменные. И все же палеоботанические документы о многом умалчивают. Мало того, что ископаемые растения доходят до нас в разобранном виде. Неполнота геологической летописи проявляется и в другом.
Мы уже рассказывали о том, что смена флоры различного типа происходит как‑то внезапно. В отдельные моменты истории непонятно, откуда берутся, причем сразу в большом количестве, совершенно новые группы растений. Потом они внезапно исчезают, оставляя немногочисленных доживающих потомков. Так было с псил офитовой флорой девона, так появилась и исчезла мезозойская флора с ее гинкговыми, цикадовыми, чекановскиевыми и беннеттитами, так вышли на сцену окружающие нас покрытосеменные. Внезапное появление новых растений и целой флоры в геологическом разрезе ‑ не свидетельство их возникновения именно в это время. Они жили и раньше (долго ли ‑ неизвестно), а мы видим лишь следы их быстрого расселения. Проникнуть в кузницу новых форм палеоботанике пока не удается, разве что в исключительных случаях. Но и тогда нельзя уверенно говорить о конкретной причине формообразования.
Поэтому на многие (хотя и далеко не на все) вопросы, касающиеся механизма эволюции растительного покрова и самих растений, приходится отвечать ботанике. Конечно, действительный ход событий могут показать лишь палеоботанические наблюдения.
Фон или движущая сила?
У колыбели каждого растения стоят злые и добрые волшебники. Одни предрекают жизненные невзгоды, другие оделяют свойствами, позволяющими побеждать врагов. Всю свою жизнь растение проводит в борьбе за воду, свет, пространство, в борьбе с болезнями, паразитами и травоядными животными. С точки зрения растения, корова ‑ беспощадный хищник.
Значение внешней среды в жизни растения умалить невозможно. Поэтому вполне понятны симпатии большинства исследователей к учению Дарвина, который считал, что именно во внешней среде надо видеть движущую силу эволюционного развития. Важнейший довод в пользу этой концепции ‑ приспособляемость организмов к различным условиям среды. Однако в науке вопросы не решаются большинством голосов. Многие биологи относятся к учению Дарвина с изрядной долей скепсиса, и это связано не с их плохим характером, а с тем, что не на все вопросы это учение отвечает достаточно убедительно. Говоря словами Э. Синнота, еще "в последней! десятилетии XIX в. энтузиазм, вызванный идеей эволюции, уступил место более здравому представлению, что эта теория не разрешает всех биологических проблем". Таких проблем немало, но мы задержимся лишь на тех из них, с которыми имеет дело палеоботаник. Только не надо думать, что автор хочет поставить под сомнение сущность эволюционного учения. Речь идет, наоборот, о возможных объяснениях с его позиций некоторых фактов из истории растительного мира.
В десятой главе мы вкратце коснулись заключительной фазы в развитии наземных растений. Примерно полтора‑два миллиона лет назад на обширные территории Земли начали наступать ледники. Подготовка к оледенению, видимо, началась довольно давно, где‑то в недрах мезозоя, когда после позднетриасового "пантропического" климата на нашей планете стала все резче и резче проявляться климатическая дифференциация. И все же, судя по всему, оледенение разразилось достаточно внезапно. Это было грандиозное событие в жизни наземных растений. Естественно ожидать, что такие кардинальные изменения условий должны вызвать и соответствующую эволюционную вспышку, но никаких следов какой‑либо вспышки не видно. За четверти чный период и предшествующие 10‑15 млн. лет мы не можем отметить появления сколько‑нибудь крупных групп растений, например порядков или семейств. Больше того, в неогеновых отложениях почти не встречаются роды, неизвестные в современной флоре, хотя географическое распределение всех родов было тогда иным, чем сейчас. Таким образом, неогеновая и четвертичная история растительного мира как будто дает в рапоряжение противников дарвинизма серьезный аргумент. Где же оно здесь, это влияние внешней среды?
Следующий вопрос задает верхнепермская флора Западной Европы. Мы уже говорили, что вся пермская история западноевропейской флоры проходила под знаком прогрессирующего иссушения климата. Не надо доказывать, что большая или меньшая сухость климата ‑ очень важный фактор внешней среды, на который должна откликнуться эволюция растений. В действительности верхпеперм‑ская флора Западной Европы составлена растениями, которые известны и в более древних отложениях, но играют там подчиненную роль. Этот факт как будто должен порадовать сторонников "теории преадаптации".
В основе этой теории лежит представление о естественном отборе, который играет роль не созидающего фактора, а своеобразного фильтра, через который на территорию с изменившимися внешними условиями проникают из окружающих мест лишь те растения (или животные), которые к ним уже приспособлены (адаптированы) по той или иной причине. Другими словами, под полог вновь образованного елового леса проникнут лишь теневыносливые растения, на ставшей засоленной почве удержатся лишь те растения, которые не погибают от избытка соли. Отсюда вывод, что не среда создает признаки и привычки организмов, а организмы подыскивают среду, подходящую к их признакам и привычкам. Что‑то похожее происходит с врачами в стране, давно освободившейся от черной оспы, если в ней неожиданно вспыхнула эпидемия. Редкие и, вроде бы, ненужные специалисты оказываются в центре внимания, становятся незаменимыми. Этим рассуждениям нельзя отказать в логичности, а анализ ископаемой флоры засушливых областей как будто подтверждает теорию преадаптации.
Этим не исчерпываются "антидарвинистские" палеоботанические документы. Во многом неясны источники многочисленных случаев параллелизма у неродственных растений, живущих далеко друг от друга и в достаточно различных условиях. Таково сходство гондванской буриадии и хвойных Северного полушария. Не менее показательно, что листья типа тениоптерис появились у самых разных групп, не связанных друг с другом ни временем, ни местом, ни (как будто) условиями жизни.
Обращает на себя внимание неравномерное развитие разных групп в одних и тех же условиях, в одном и том же месте. Одни группы прогрессируют, дают множество видов, тогда как другие долго живут на том же фоне, обходясь без существенных нововведений. Каверзные вопросы задают и древнейшие растения. Мы уже говорили об их внезапном появлении и необычайно быстром прогрессе, хотя никаких выдающихся стимулирующих событий во внешних условиях в то время, видимо, не было. Наконец, последовательность фаз складчатости и горообразования, менявших лик континентов, не совпадает с важнейшими этапами в эволюции наземных растений.
Как же можно с позиций дарвинизма истолковать эти и многие другие палеоботанические факты? Неужели действительно внешняя среда лишь фон, на котором развертываются эволюционные процессы?
Почему "или..., или..."?
Прежде чем отвечать на поставленные вопросы, четко разграничим два понятия: происхождение организмов и их появление в геологическом разрезе. Оба понятия не столь тесно связаны друг с другом, как кажется на первый взгляд. При рассмотрении общих вопросов эволюции палеонтологи любят списывать отсутствие документов на неполноту геологической летописи. Эта неполнота действительно настолько значительна, что известный немецкий ботаник К. Гебель сравнил палеоботанические свидетельства с жалкими обрывками растерянной библиотеки. Однако оснований для такого пессимизма, пожалуй, все же нет. Выпадение страниц из геологической летописи ‑ закономерный процесс, и их отсутствие само по себе достаточно красноречиво.
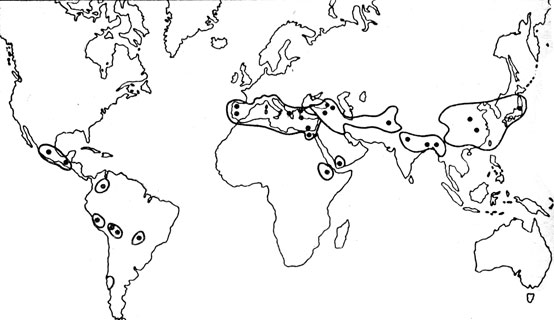
Вот один пример. Изучая закономерности распределения культурных растений, Н. И. Вавилов обратил внимание на то, что на Земном шаре есть места с огромным разнообразием сортов того или иного растения. Так, в Эфиопии на площади не более, чем полмиллиона гектаров, было обнаружено больше видов и разновидностей пшеницы, чем во всех остальных местах Земного шара вместе взятых. Работы Вавилова и его учеников показали, что подобные концентрации не случайны. Они указывают нам на центры формообразования, некие генераторы новых видов и разновидностей. Больше того, Вавилов пишет о фактах "изумительной приуроченности процесса основного формообразования географически к чрезвычайно малым пространствам". Выяснилось, что современные очаги сортовых богатств приурочены к горным районам Азии, к Гималаям и их отрогам, к горным системам Северо‑Восточной Африки, Южной Европы (Пиренеи, Апеннины, Балканы), Кордильерам, южным отрогам Скалистых гор (рис. 47). В Старом Свете речь идет о полосе между 20 и 40° с. ш. Эти горные области граничат с пустынями Центральной Азии, Сахарой и по разнообразию климата и почв представляют оптимальные условия для проявления формообразовательного процесса. По количеству осадков, по температурным различиям, по почвенным типам здесь выражены градации до крайних вариантов. Важно и то, что "географическая локализация видообразования культурных растений в значительной мере совпадает с локализацией общего видообразовательного процесса, свойственного флорам всего Земного шара".
Основные центры происхождения культурных растений (по Н. И. Вавилову)
Из сказанного следует очень важный для палеоботаники вывод. Поскольку подавляющая часть древних растений, известных палеоботаникам, населяла болота и низины, а отнюдь не предгорья, есть мало шансов встретить "документы" о формообразовании в геологическом прошлом. Отсюда следует другой вывод: смена флоры, часто очень резкая, которую мы видим в разрезе, не имеет непосредственного отношения к видообразованию. Это уже следующая стадия в развитии растительного покрова, а именно ‑ расселение в новые условия флоры, сформировавшейся за кулисами наблюдаемых нами событий.
О таком механизме развития растительного покрова в геологическом прошлом писали Ч. Дарвин, английский палеоботаник Д. Г. Скотт (в 20‑х годах), но полнее всего он был вскрыт в работах нашего крупнейшего палеоботаника А. Н. Криштофовича. Поведение растений при резком изменении внешних условий (оледенение, аридизация) А. Н. Криштофович сравнивал с паническим бегством кошки, взбирающейся на дерево при виде собаки. Действительно, растениям, хорошо приспособленным к внешней среде, нужно время для перестроек, если эта среда меняется. Быстрые изменения климата и почвы приводят растение к гибели. Для эволюции нужны гомеопатические дозы направленных изменений.
Представление о том, что основное формообразование совершается в приподнятых участках суши и поэтому выпадает из нашего поля зрения, было широко взято на вооружение палеоботаниками. Сейчас на это явление ссылаются каждый раз, когда видят внезапное появление групп растений и целой флоры. Но как же тогда быть с раннедевонскими событиями в мире наземных растений? Неужели и здесь внезапное появление в разрезе богатой ископаемой флоры зависит от той же причины? Нет, едва ли. Очень трудно, почти невозможно отказаться от представления, что колыбель наземных растений ‑ побережье морей или других водоемов. Предками наземных растений были водоросли. Таково единодушное мнение исследователей, и подвергать его сомнению пока нет оснований. Предполагать же, что водоросли спустились в низины, т. е. в область формирования флороносных толщ, начав свой путь в предгорьях, по меньшей мере смело.
Причину внезапного появления богатства раннедевонской флоры, видимо, надо искать в другом.
Растительность не всегда сплошь покрывала континенты. Когда‑то суша была вовсе безжизненной. Первые растения стали проникать в глубь ее по отдельным увлажненным участкам. В этих условиях на континентах не было настоящего почвообразования, не регулировался сток вод. Возможно, что по этой причине все континентальное и прибрежно‑морское осадконакопление шло совсем не так, как в последние геологические эпохи. Иными словами, другими не достаточно устойчивыми были и "кладбища" древнейших растений. По мере эволюции наземные растения становились способными к более полному заселению суши, развитие корневой системы позволяло им прочнее удерживаться на субстрате, а значит и закреплять его.
Даже небольшое приобретение в этом отношении могло резко изменить всю ситуацию. Образуется единая связанная цепь: больше растений, крепче они сидят ‑ лучше регулируется сток ‑ меньше размыв склонов ‑ меньше повреждений в растительном покрове ‑ больше растений и т. д. Процесс нарастал лавинообразно, в очень короткие сроки. Суша быстро приобрела обжитой вид, стали формироваться вполне сносные "кладбища" растений, но углеоб‑разование еще не начиналось. Позже те же процессы, развиваясь и углубляясь, настолько изменили характер стока вод и почвообразования, что стало возможным формирование болотистых низин, в которых стали накапливаться угли. Кстати, последние появляются в геологическом разрезе также очень неожиданно. Девонские угли редки, зато в нижнекаменноугольных отложениях их уже много во многих местах Земного шара.
Экспансия на незанятые участки суши при незначительной конкуренции и богатом воздействии факторов, каждый из которых был в диковинку выходцам из воды, ‑ вот, по‑видимому, тот источник, которому обязан девонский растительный мир своей сверхскоростной эволюцией.
На несколько каверзных палеоботанических вопросов мы как будто подобрали более или менее сносные ответы. Можно объяснить и неодинаковые темпы эволюции разных растений в одних и тех же условиях: во‑первых, изменения не обязательно захватывают все компоненты среды в равной мере; во‑вторых, разные растения обладают не одинаковой мерой эволюционной пластичности.
Труднее объяснить с точки зрения дарвиновской теории естественного отбора многочисленные случаи параллелизмов, о которых уже много говорилось в девятой главе. Во всяком случае, здесь огромную роль должны играть сложные генетические механизмы. Нельзя исключать и возможные проявления направленного развития, происходящего при известной автономии от внешней среды.
Вопрос о происхождении параллелизмов не может быть окончательно решен, пока не выяснено приспособительное значение параллельно появляющихся признаков. До этого еще слишком далеко, и мы вынуждены занимать в отношении влияния среды довольно примирительную позицию, не ставя решительно вопрос "или..., или...", помня о том, что большинство кардинальных вопросов науки находило решение тогда, когда исследователи принимали во внимание "и..., и...". Точно так же нельзя полностью отвергать и всю концепцию преадаптации. Полемизируя с ее сторонниками, палеонтолог Помпецкий справедливо заметил, что ихтиозавры едва ли перешли к жизни в море по той причине, что у них конечности стали преображаться в ласты.
Однако, когда мы имеем дело с быстрыми и сильными изменениями среды, мы видим, что из окружающих мест на эти участки проникают лишь те, кто может жить в этих ставших крайними условиях. Здесь идет не столько изменение, сколько переселение организмов. Например, в третичное (особенно раннетретичное, т. е. палеогеновое) время на Земле, видимо, не было широких областей с арктическим климатом. По тем временам тундры и лесотундры, очевидно, были именно такой крайней и нетипичной средой. Но четвертичное оледенение резко изменило картину. И в наши дни холодный климат господствует на обширных территориях. Последние заселялись, по‑видимому, главным образом теми растениями, которые уже научились обходиться без большого количества тепла.
Ускоряется ли прогресс?
В числе основных проблем биологии, к решению которых палеоботаника (и вообще палеонтология) имеет прямое отношение, стоит проблема эволюционного прогресса. Среди зарубежных ученых есть немало таких, которые отрицают органический прогресс или считают его незначительным. Даже знаменитый дарвинист Томас Гексли, страстный проповедник эволюционного учения, писал, что "суммарный итог" развития органического мира "изумительно незначителен". Этой точке зрения можно противопоставить высказывание палеоботаника Г. П. Радченко, который довольно категорично пишет: "Прогрессивная эволюция органического мира выражается в повышении уровня организации животных и растений на протяжении всей истории биосферы и имеет ясно выраженную тенденцию к постепенному убыстрению этого процесса". А вот один из крупнейших современных генетиков, шведский ученый А. Мюнтцинг пишет: "... В настоящее время едва ли продолжается процесс эволюции в направлении возрастания сложности организации". Мы ограничимся этими взятыми наудачу высказываниями, поскольку привести все точки зрения, изложенные в литературе, совершенно невозможно. Их рассмотрению посвящены обстоятельные научные статьи и целые монографии.
Вопрос об эволюционном прогрессе при всей его кажущейся простоте (неужели развитие от амебы до человека не свидетельство прогресса?) не может считаться окончательно решенным. Достаточно сказать, что генетические механизмы удивительно сходны у огромного большинства организмов. Поэтому генетики, изучая наследственность дрозофилы, могут применять многие полученные закономерности к человеку. Таким образом, по такому важнейшему признаку всего живого, как наследственность, мы не в состоянии указать на сколько‑нибудь значительный прогресс в течение последнего полумиллиарда лет.
Трудность определения прогресса связана еще и с другим обстоятельством. Гигантские динозавры, если бы они были наделены разумом и могли ознакомиться с остатками своих предков, наверное, сочли бы себя достаточно прогрессивными. Тем не менее динозавры, некогда господствовавшие на суше, на море и в воздухе вымерли, а их далекий примитивный предок кистеперая рыба латимерия дожила до наших дней. Оценка прогресса с точки зрения высоты и сложности организма ‑ дело рискованное.
Часто о прогрессивности животного судят по тому, находилось ли оно на филогенетической дороге, ведущей к человеку. Мы сами стали в глазах многих биологов мерилом прогресса. Это, конечно, довольно шаткий критерий, ибо судьба человечества в масштабах геологического времени неизвестна. Кто знает, может быть, прав окажется Реми Шовен. Он, шутя, пишет о том, что "лет этак миллионов через сто какой‑нибудь шестиногий эрудит напишет, что судя по весьма древним источникам, некая обезьяна совершенно внезапно превратилась в другое существо и это новое существо настроило невесть чего, каких‑то гор из камня, а затем молниеносно исчезло ..."
Тем не менее полностью отрицать прогресс трудно. Вслед за большинством наших исследователей мы примем, что прогресс по крайней мере весьма вероятен.
Раз так, то интересно определить меру прогресса растений в разные геологические периоды. Здесь, очевидно, нужно вести оценку по двум показателям: сначала по появлению тех или иных признаков, а затем по их комбинациям. И тут мы сталкиваемся с очень интересными фактами. Расчленим известные нам наземные растения на отдельные признаки и посмотрим, в какой последовательности они появлялись в геологической истории. Оказывается, что еще в раннем девоне наметились основные признаки внутренней структуры. В конце девона внутренняя структура достигла уже очень высокой дифференциации. С этого времени и до мезозоя в ней не отмечается серьезных усовершенствований. Правда, во второй половине мезозоя появились покрытосеменные, важным приобретением которых были сосуды. Девонские растения преуспели не только в анатомическом строении. У них уже были семезачатки. До появления покрытосеменных семезачатки не претерпели существенных усложнений. У покрытосеменных семезачатки укрыты сросшимися покровами, усложнился и процесс оплодотворения.
Что касается строения листвы, то и здесь основные успехи также принадлежат девонским растениям. Правда, у них еще нет листьев, в которых жилки располагаются перисто, а также листьев с сетчатым жилкованием. Однако все это появилось уже к середине каменноугольного периода.
Большим достижением было приобретение покрытосеменными травянистого облика. О преимуществах травянистых (особенно однолетних) растений уже шла речь выше. Когда появились на Земле такие покрытосеменные, мы точно не знаем, но возможно, что они были уже во второй половине мелового периода.
К сожалению, сделать такую же оценку не отдельных признаков, а целых организмов, мы почти не можем из‑за неполноты тех растительных остатков, которые попадают в руки палеоботаников. Однако и сказанного достаточно, чтобы предположить, что в истории наземных растений основной прогресс совершался в два этапа. Первый этап падает на девонский период, второй ‑ на середину мезозоя. В последние геологические эпохи растения, занимаясь мелкими усовергаенствованиямрт, не достигли чего‑нибудь необычного.
Таким образом, из трех точек зрения на прогресс, названных в начале этого раздела, приходится склоняться к точке зрения, высказанной генетиком Мюнтцингом. Мы не можем отрицать прогресс в растительном мире, по крайней мере при сегодняшнем состоянии наших знаний. Он явно затронул внешний облик растений и позволил им захватить огромные территории. Но говорить о каком‑либо решительном ускорении прогресса растений все же трудно. Скорее, наоборот, сейчас мы являемся свидетелями спокойной их эволюции, не ведущей к существенному усложнению организации. Это вполне понятно в условиях суши, обильно заселенной прекрасно приспособленными организмами, опытными конкурентами в борьбе за существование.
Если дружно мы навалимся...
В том, что разобранные выше вопросы интересны, по‑видимому, нет сомнений. Они не только интересны, но и важны, и об их значении мы будем говорить в следующей главе. Однако до решения их еще далеко. Впрочем, это судьба всех кардинальных вопросов биологии. Решение оттягивается тем, что для пего нужны совместные усилия специалистов в разных областях знаний: ботаников, палеоботаников, геологов, генетиков. Как теперь модно говорить, нужен комплексный подход к решению этих проблем.
Действительно, эволюция ископаемых растений не может изучаться в отрыве от современной биологии. Уже установлено множество любопытных биологических механизмов, эволюционное значение которых по‑настоящему не оценено. Например, лишь немногие палеоботаники учитывают в своих эволюционных построениях неотенические явления. Речь идет о способности организмов приступать к размножению до завершения индивидуального развития, иногда даже в зародышевом состоянии. В работах А. Л. Тахтаджяна, И. Т. Васильченко и других исследователей отчетливо показано, какую огромную роль сыграла неотения в эволюции покрытосеменных. А. Л. Тахтаджян полагает, что именно с неотенией связано появление и травянистого облика у многих покрытосеменных, и самой покрытосемянности. О явлениях неотении в других группах растений мы знаем очень мало.
Вот другой пример. В работах, посвященных общим проблемам палеоботаники, почти не встретишь слова "апомиксис". Этим термином обозначают случаи замещения полового размножения бесполым. Такое бесполое размножение может быть как вегетативным (посредством отводков, клубней, луковиц и т. д.), так и с образованием внешне обычных семян, которые на самом деле развиваются без оплодотворения. Растения‑апомикты имеют ряд преимуществ, иногда решающих. Благодаря этому явлению, хорошо приспособленный к данным условиям вид может быстро воспроизводиться в массовых количествах при совершенно одинаковой и притом оптимальной генетической структуре. При половом процессе изменчивость, часто вредная, значительно больше. Апомикты неизменно доставляют много хлопот систематикам, так как вместо широкого диапазона изменчивости у них образуется множество постоянных типов, каждый из которых мало отличается друг от друга. К апомиктам, кстати, относится ястребинка, которая давала несколько другое расщепление признаков у гибридов, нежели горох, чем и заставила Г. Менделя сомневаться во всеобщности выведенных им закономерностей. Апомик‑тичные растения ‑ сущее наказание для систематика, который, видя выдержанность изменений (хотя и мелких), описывает все новые и новые виды, в действительности имея дело с одними тем же видом. Может быть, учет таких явлений палеоботаниками заставит их быть более осторожными и снизит количество описываемых в литературе новых видов ископаемых растений.
Число подобных примеров, когда генетика может прийти на помощь в решении палеоботанических проблем, можно легко увеличить. Опыт показывает, что союз генетики и палеозоологии очень плодотворен (вспомним интереснейшие работы Дж. Г. Симпсона). Почему же науке об ископаемых растениях быть сиротой?
Точно такие же тесные связи могут быть установлены палеоботаникой с палеогеографией, учением о накоплении осадочных полезных ископаемых, географией растений и многими другими дисциплинами. Но, к сожалению, эти связи слабы и не постоянны. От этого страдают и палеоботаника, и перечисленные области знаний. В чем же дело, что мешает все расставить по своим местам, что мешает установлению всех этих плодотворных связей? Что это ‑ вина или беда палеоботаников? С этим вопросом мы попробуем разобраться в следующей главе.
|
К содержанию: Мейнен: ИЗ ИСТОРИИ РАСТИТЕЛЬНЫХ ДИНАСТИЙ
Смотрите также:
Необратимость эволюции и прогрессивный ход эволюции.
Когда появились первые деревья. Каменноугольный период, или...
Что такое палеоботаника или ботаническая палеонтология...
БИОЛОГИЯ. Пособие по биологии, автор Мамонтов
Ангиосперм. Появление и распространение покрытосеменных...
Эволюция растений. Поздний палеозой — ранний мезозой...
Основные пути эволюции наземных растений